 中文网站
中文网站
Under PCR-reaktionen opstår der ofte nogle interfererende faktorer.
På grund af PCR's meget høje følsomhed anses kontaminering for at være en af de vigtigste faktorer, der påvirker PCR-resultater, og kan give falsk positive resultater.
Lige så kritiske er de forskellige kilder, der fører til falsk negative resultater. Hvis en eller flere væsentlige dele af PCR-blandingen eller selve amplifikationsreaktionen hæmmes eller forstyrres, kan den diagnostiske analyse blive hindret. Dette kan føre til reduceret effektivitet og endda falsk negative resultater.
Ud over hæmning kan der forekomme tab af målnukleinsyrens integritet på grund af forsendelses- og/eller opbevaringsforhold før prøveforberedelse. Især høje temperaturer eller utilstrækkelig opbevaring kan føre til beskadigelse af celler og nukleinsyrer. Celle- og vævsfiksering og paraffinindlejring er velkendte årsager til DNA-fragmentering og et vedvarende problem (se figur 1 og 2). I disse tilfælde vil selv optimal isolering og oprensning ikke hjælpe.
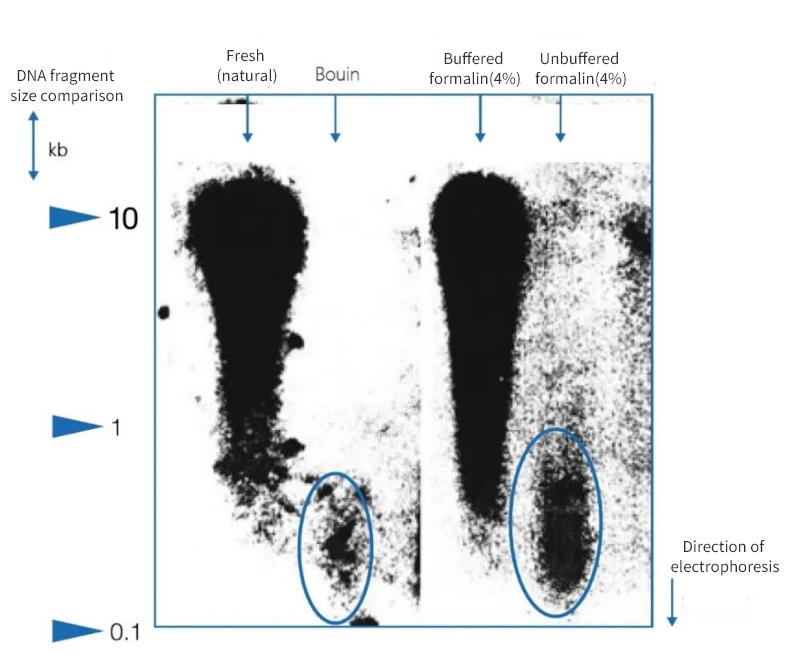
Figur 1 | Effekt af immobilisering på DNA-integritet
Agarosegelelektroforese viste, at kvaliteten af DNA isoleret fra paraffinsektioner fra obduktioner varierede betydeligt. DNA med forskellige gennemsnitlige fragmentlængder var til stede i ekstrakterne afhængigt af fikseringsmetoden. DNA blev kun bevaret, når det blev fikseret i native frosne prøver og i bufferet neutral formalin. Brugen af et stærkt surt Bouin-fiksativ eller ubufferet, myresyreholdig formalin resulterede i et betydeligt tab af DNA. Den resterende fraktion er stærkt fragmenteret.
Til venstre er fragmenternes længde udtrykt i kilobasepar (kbp)
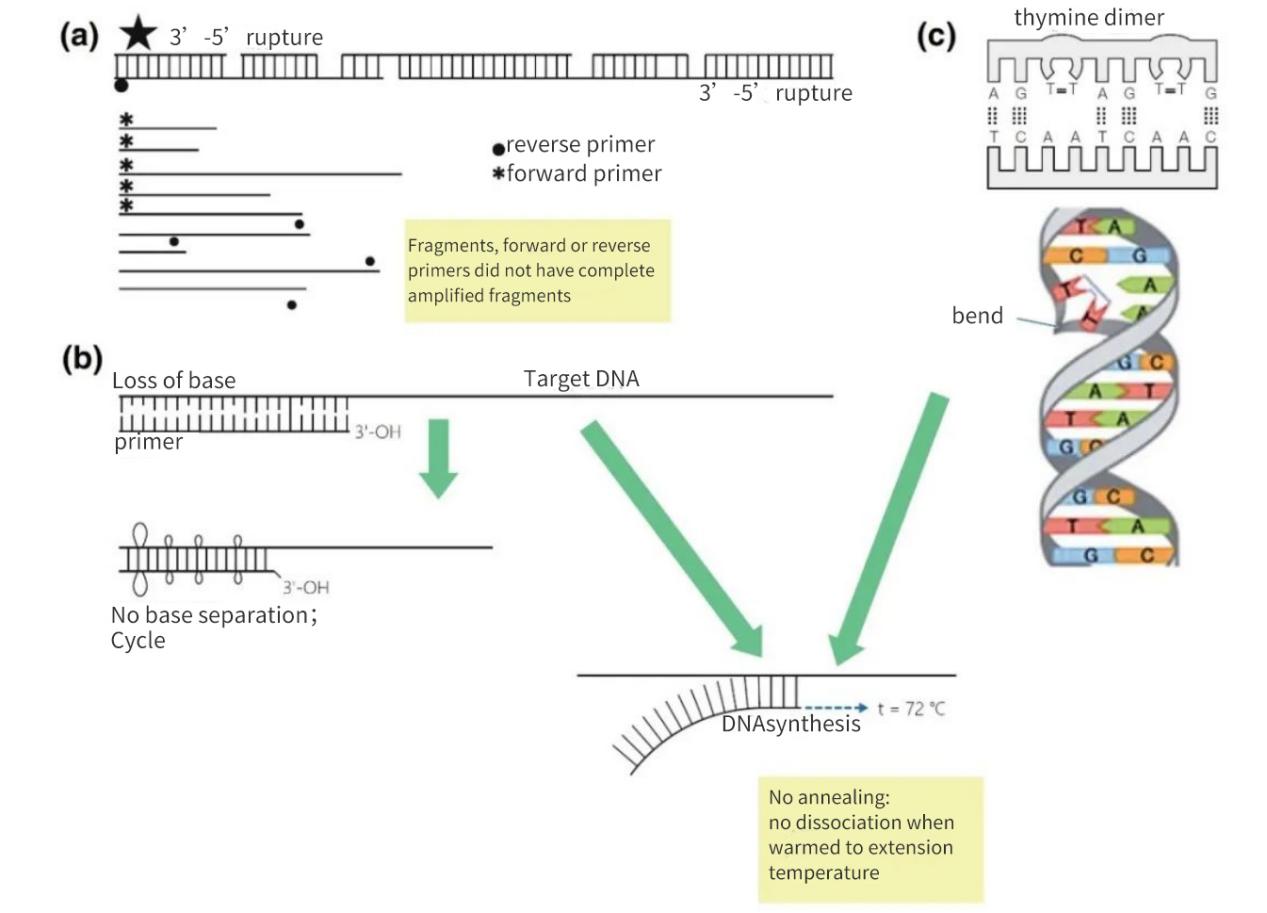
Figur 2 | Tab af integritet af nukleinsyremål
(a) Et 3′-5′ mellemrum på begge strenge vil resultere i et brud i mål-DNA'et. Syntese af DNA vil stadig forekomme på det lille fragment. Hvis der mangler et primer-annealingssted på DNA-fragmentet, forekommer der dog kun lineær amplifikation. I det mest gunstige tilfælde kan fragmenterne mætte hinanden igen, men udbytterne vil være små og under detektionsniveauerne.
(b) Tab af baser, primært på grund af depurinering og dannelse af thymidin-dimerer, fører til et fald i antallet af H-bindinger og et fald i Tm. Under den forlængede opvarmningsfase vil primerne smelte væk fra matrix-DNA'et og vil ikke anneale selv under mindre stringente betingelser.
(c) Tilstødende thyminbaser danner en TT-dimer.
Et andet almindeligt problem, der ofte opstår i forbindelse med molekylær diagnostik, er den mindre optimale frigivelse af målnukleinsyrer sammenlignet med phenol-chloroform-ekstraktion. I ekstreme tilfælde kan dette være forbundet med falsk negative resultater. Der kan spares meget tid ved kogende lysis eller enzymatisk nedbrydning af cellerester, men denne metode resulterer ofte i lav PCR-følsomhed på grund af utilstrækkelig frigivelse af nukleinsyrer.
Hæmning af polymeraseaktivitet under amplifikation
Generelt bruges hæmning som et begreb til at beskrive alle faktorer, der fører til suboptimale PCR-resultater. I en strengt biokemisk forstand er hæmning begrænset til enzymets aktivitet, dvs. den reducerer eller forhindrer substrat-produkt-omdannelse gennem interaktion med det aktive sted i DNA-polymerasen eller dens cofaktor (f.eks. Mg2+ for Taq DNA-polymerase).
Komponenter i prøven eller forskellige buffere og ekstrakter, der indeholder reagenser, kan direkte hæmme enzymet eller fange dets cofaktorer (f.eks. EDTA), hvorved polymerasen inaktiveres og dermed føre til nedsatte eller falsk negative PCR-resultater.
Mange interaktioner mellem reaktionskomponenter og målholdige nukleinsyrer betegnes dog også som 'PCR-hæmmere'. Når cellens integritet forstyrres af isolering, og nukleinsyren frigives, kan der forekomme interaktioner mellem prøven og dens omgivende opløsning og faste fase. For eksempel kan 'fjernere' binde enkelt- eller dobbeltstrenget DNA gennem ikke-kovalente interaktioner og forstyrre isolering og oprensning ved at reducere antallet af mål, der til sidst når PCR-reaktionsbeholderen.
Generelt findes PCR-hæmmere i de fleste kropsvæsker og reagenser, der anvendes til kliniske diagnostiske tests (urinstof i urin, hæmoglobin og heparin i blod), kosttilskud (organiske komponenter, glykogen, fedt, Ca2+-ioner) og komponenter i miljøet (fenoler, tungmetaller).
| Hæmmere | Kilde |
| Calciumioner | Mælk, knoglevæv |
| Kollagen | Væv |
| Galdesalte | Afføring |
| Hæmoglobin | I blod |
| Hæmoglobin | Blodprøver |
| Humussyre | Jord, plante |
| Blod | Blod |
| Laktoferrin | Blod |
| (Europæisk) melanin | Hud, hår |
| Myoglobin | Muskelvæv |
| Polysakkarider | Plante, afføring |
| Protease | Mælk |
| Urea | Urin |
| Mukopolysaccharid | Brusk, slimhinder |
| Lignin, cellulose | Planter |
Mere udbredte PCR-hæmmere kan findes i bakterier og eukaryote celler, ikke-mål-DNA, DNA-bindende makromolekyler i vævsmatricer og laboratorieudstyr såsom handsker og plastik. Oprensning af nukleinsyrer under eller efter ekstraktion er den foretrukne metode til fjernelse af PCR-hæmmere.
I dag kan forskellige automatiserede ekstraktionsudstyr erstatte mange manuelle protokoller, men 100% genvinding og/eller oprensning af mål er aldrig blevet opnået. Potentielle inhibitorer kan stadig være til stede i de oprensede nukleinsyrer eller kan allerede have virket. Der findes forskellige strategier til at reducere inhibitorernes virkning. Valget af den passende polymerase kan have en betydelig indflydelse på inhibitoraktiviteten. Andre dokumenterede metoder til at reducere PCR-hæmning er at øge polymerasekoncentrationen eller anvende tilsætningsstoffer såsom BSA.
Hæmning af PCR-reaktioner kan påvises ved brug af intern proceskvalitetskontrol (IPC).
Der skal udvises omhu med at fjerne alle reagenser og andre opløsninger i ekstraktionskittet, såsom ethanol, EDTA, CETAB, LiCl, GuSCN, SDS, isopropanol og phenol, fra nukleinsyreisolatet ved en grundig vask. Afhængigt af deres koncentration kan de aktivere eller hæmme PCR.
Udsendelsestidspunkt: 19. maj 2023
 Privatlivsindstillinger
Privatlivsindstillinger